
Sewall Wright 1889-ben született Massachusettsben. Illinois-ban tanult, ahol édesapja közgazdaságtant tanított. Miután az Illinois Egyetemen mesterfokozatot szerzett biológiából, a Harvard Egyetemen szerzett PhD-t a tengerimalacok bőrszíne öröklődésének témájában. 1915 és 1925 között tovább folytatta tengerimalacokkal kapcsolatos kutatásait Washingtonban. Később a Chicagói Egyetemen dolgozott. Fisher 1922-es cikkének hatására Wright 1925-ben hosszú cikket írt Evolution in Mendelian populations címmel, amely végül 1931-ben jelent meg, és amelyben egy olyan modellt vizsgált, amely implicit módon Fisher 1930-ban megjelent, The General Theory of Natural Selection című könyvében is megjelent. Akárcsak a Hardy-Weinberg-törvény esetén, ez a modell szintén azt az esetet tekinti, amikor csak két lehetséges allél - A és a - van egy lókuszon, de a populációról nem tesszük fel, hogy végtelen nagyságú. A cél az volt, hogy kiderüljön: ennek a feltételnek az elhagyása milyen hatással van a populáció genetikai összetételére.
tanított. Miután az Illinois Egyetemen mesterfokozatot szerzett biológiából, a Harvard Egyetemen szerzett PhD-t a tengerimalacok bőrszíne öröklődésének témájában. 1915 és 1925 között tovább folytatta tengerimalacokkal kapcsolatos kutatásait Washingtonban. Később a Chicagói Egyetemen dolgozott. Fisher 1922-es cikkének hatására Wright 1925-ben hosszú cikket írt Evolution in Mendelian populations címmel, amely végül 1931-ben jelent meg, és amelyben egy olyan modellt vizsgált, amely implicit módon Fisher 1930-ban megjelent, The General Theory of Natural Selection című könyvében is megjelent. Akárcsak a Hardy-Weinberg-törvény esetén, ez a modell szintén azt az esetet tekinti, amikor csak két lehetséges allél - A és a - van egy lókuszon, de a populációról nem tesszük fel, hogy végtelen nagyságú. A cél az volt, hogy kiderüljön: ennek a feltételnek az elhagyása milyen hatással van a populáció genetikai összetételére.
Jelölje N az összes egyed számát, erről feltesszük, hogy minden generációban megegyezik. Minden egyed két alléllel rendelkezik, vagyis összesen 2N allél van a populációban minden generációban. Azt is feltesszük, hogy a párválasztás véletlenszerűen történik. Ha az n-edik generációban i darab A allél és 2N-i darab a allél van, akkor az n+1-edik generációban egy véletlenszerűen kiválasztott allél i/(2N) valószínűséggel lesz A, 1-i/(2N) valószínűséggel pedig a típusú. Tehát annak a valószínűségét, hogy az n+1-edik generációban az A típusú allélok száma j-vel egyenlő, a következő képlet adja meg:

Jelölje az Xn véletlen változó az A típusú allélok számát.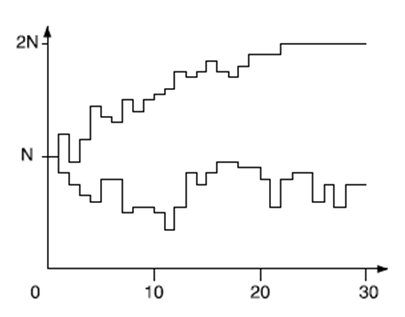
Két szimuláció, Xn változására 30 generáción keresztül, N=20 és X0=10 esetén
Megmutatható, hogy ha tudjuk, hogy Xn=i, akkor Xn+1 várható értéke szintén i; ez a Hardy-Weinberg-törvényre emlékeztet, ahol az A allél gyakorisága változatlan volt a generációkon át. Most azonban előfordulhat, hogy X0=i kezdeti feltételből indulva (ahol 0<i<2N) bizonyos számú generáció után az Xn=0 esemény következik be. Ebben az esetben minden allél a típusú lenne, és Xn az összes további generációban 0 lenne. Természetesen a fordított eset is bekövetkezhet. Azaz, ha feltesszük, hogy a populáció végtelen nagyságú, mint a Hardy-Weinberg-modell esetében, akkor a két allél nem tűnhet el, mivel arányuk konstans marad, ha viszont figyelembe vesszük a populáció véges méretét, mint a Fisher-Wright-modellben, akkor a két allél aránya változik, és az egyik allél eltűnik.
X0=i-ből indulva egyszerűen kiszámíthatjuk annak a valószínűségét, hogy a populáció az X=0 fixpontba kerül. Jelöljük ezt a valószínűséget Qi-vel, természetesen Q0=1 és Q2N=0. Továbbá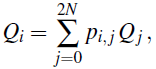
mivel pi,jQj annak a valószínűsége, hogy az X=0 fix állapotba kerülünk az X0=i kezdeti állapotból indulva, az X1=j állapoton keresztül. Mivel a pi,j-ket i-re összegezve 1-et kapunk, adódik, hogy Qi=1-i/(2N) az előbbi egyenletünk megoldása a fenti peremfeltételekkel. Ezért annak a valószínűsége, hogy egy N méretű populációban i db A típusú allélből indulva egy csak a típusú allélt tartalmazó populációhoz jutunk, 1-i/(2N)-nel egyenlő, míg annak a valószínűsége, hogy egy csak A alléleket tartalmazó populációhoz jutunk, i/(2N). Wright megmutatta, hogy a fixállapot eléréséhez szükséges generációszám 2N nagyságrendű. Egy több milliós populáció esetén ez olyan sok ideig tartana, hogy az allélek gyakoriságát majdnem konstansnak tekinthetjük, akárcsak a Hardy-Weinberg-modell esetén.
1922-ben Fisher már megpróbálta megbecsülni a fixáció rátáját, de elnézett egy 2-es szorzót. Fisher és Wright eltérő véleményen volt a populációk tipikus méretét illetően. Az evolúció elméletének szempontjából Wright munkája azt sugallta, hogy kis méretű populációk esetén a véletlen genetikai sodródás a fajok eredetének mechanizmusa lehet. A fajok besorolásán dolgozó biológusok észrevették, hogy a fajok és alfajok közti különbségekre sokszor nem ad megfelelő magyarázatot a természetes szelekció. Az 1940-es és 50-es években ezt a felvetést erősen ellenezte Fisher és kollégája, E. B. Ford, mivel úgy gondolták, hogy a véletlen genetikai sodródás elhanyagolható a természetes szelekcióhoz képest. Elsősorban arra a tanulmányukra hivatkoztak, amelyben egy Oxfordhoz közeli, kicsi, izolált molypopuláció (Panaxia dominula) génarányainak fluktuációját vizsgálták. Ebben a populációban a három különböző genotípus (gyakori homozigóta, heterozigóta, ritka homozigóta) jól megkülönböztethető sabad szemmel.
Az újabb, realisztikusabb modellek egyszerre veszik figyelembe a természetes szelekciót, a véletlen genetikai sodródást, a mutációt, a migrációt és egyéb tényezőket. A véletlen sodródás szerepét a japán Kimura hangsúlyozta ki újra.
Wright 1934-ben a Tudományos Akadémia tagja lett. Sokáig a Death Valley-beli légypopulációk (Drosophila pseudoobscura) genetik
Ajánlott bejegyzések:
A bejegyzés trackback címe:
Kommentek:
A hozzászólások a vonatkozó jogszabályok értelmében felhasználói tartalomnak minősülnek, értük a szolgáltatás technikai üzemeltetője semmilyen felelősséget nem vállal, azokat nem ellenőrzi. Kifogás esetén forduljon a blog szerkesztőjéhez. Részletek a Felhasználási feltételekben és az adatvédelmi tájékoztatóban.